
拉丁文名称:Cypripedium tibeticum King ex Hemsl.
拼音:xi zang shao lan
科 属:兰科Orchidaceae,杓兰属Cypripedium
拉丁文释义:Cypri- 就是塞浦路斯,也是维纳斯的别名(比维纳斯得名还早)。-pedium则是足、拖鞋的意思。Cypripedium就是女神的拖鞋。杓兰自然分布在北温带偏北。比如美国“大陆四十八州”中最北的明尼苏达州,其州花就是女神的拖鞋,在那么冷的地区,又是北美,这女神的拖鞋当然就是杓兰了。
国内分布:产甘肃南部、四川西部、贵州西部、云南西部和西藏东部至南部。生于海拔2300--4200米的透光林下、林缘、灌木坡地、草坡或乱石地上。本种后选模式标本产四川。
国外分布:不丹和锡金也有分布。
海 拔:2300-4200
命名来源:[Journal of the Linnean Society, Botany 29(202): 320 1892]
中国植物志:17:036
亚组中文名:大花亚组
亚组拉丁名:Subsect.Macrantha
组中文名:杓兰组
组拉丁名:Sect.Cypripedium
属中文名:杓兰属
属拉丁名:Cypripedium
亚科中文名:杓兰亚科
亚科拉丁名:Subfam.CYPRIPEDIOIDEAE Lindl.
科中文名:兰科
科拉丁名:Orchidaceae
纲中文名:单子叶植物纲
纲拉丁名:MONOCOTYLEDONEA
门中文名:被子植物门
门拉丁名:ORCHIDACEAE
形态特征:植株高15—35厘米,具粗壮、较短的根状茎。茎直立,无毛或上部近节处被短柔毛,基部具数枚鞘,鞘上方通常具3枚叶,罕有2或4枚叶。叶片椭圆形、卵状椭圆形或宽椭圆形,长8—16厘米,宽3---9厘米,先端急尖、渐尖或钝,无毛或疏被微柔毛,边缘具细缘毛。花序顶生,具1花;花苞片叶状,椭圆形至卵状披针形,长6—11厘米,宽2--5厘米,先端急尖或渐尖;花梗和子房长2--3厘米,无毛或上部偶见短柔毛;花大,俯垂,紫色、紫红色或暗栗色,通常有淡绿黄色的斑纹,花瓣上的纹理尤其清晰,唇瓣的囊口周围有白色或浅色的圈;中萼片椭圆形或卵状椭圆形,长3--6厘米,宽2.5--4厘米,先端渐尖、急尖或具短尖头,背面无毛或偶见疏微柔毛,边缘多少具细缘毛;合萼片与中萼片相似,但略短而狭,先端2浅裂;花瓣披针形或长圆状披针形,长3.5—6.5厘米,宽1.5--2.5厘米,先端渐尖或急尖,内表面基部密生短柔毛,边缘疏生细缘毛;唇瓣深囊状,近球形至椭圆形,长3.5-----6厘米,宽亦相近或略窄,外表面常皱缩,后期尤其明显,囊底有长毛;退化雄蕊卵状长圆形,长1.5—2厘米,宽8—12毫米,背面多少有龙骨状突起,基部近无柄。花期5—8月。
产地分布:产甘肃南部、四川西部、贵州西部、云南西部和西藏东部至南部。生于海拔2300--4200米的透光林下、林缘、灌木坡地、草坡或乱石地上。不丹和锡金也有分布。本种后选模式标本产四川。
参考文献:CypripediumtibeticumKingexRolfeinJ.Linn.Soc.Bot.29:320.1892;KingetPan-tl.inAnn.Bot.Gard.Calcutta8:341.Pl.447.1890;RolfeinJ.Linn.Soc.Bot.36:66.1903,etinCurtis'sBot.Mag.132:t.8070.1906;Sehhr.InFeddeRepert.Sp.Nov.Beih.4:85.1919;S.Y.HuinQuart.J.TaiwanMus.26(1--2):148.1973;S.C.ChenetY.Z.XiinProc.12thWoddOrch.Conf.144.1987;K.Y.LangetZ.H.TsiinBot.Res.Inst.Bot.6:49.1992;横断山区维管植物2:2511.1994;Z.H.TsiinSel-byanal8(1):17.1997;Cribb,GenusCypripedium204.Fig32.Pl.25,110--116.1997;Z.H.Tsi,S.C.ChenetK.Mori,WildOreh.Chinal2.Pl.2.1997;中国兰花全书115.图6--3(4).彩图6--7.1998.——CypripediumcorrugatumFranch.InJ.Bot.(Morot)8:251.1894.——CypripediumcorrugatumFranch.Var.ObesumFranch.L.C.1894.——CypripediummacranthumSw.var.Tibeticum(KingXRolfe)Kraenzl.,Orch.Gen.Sp.1:26.1897.——Cypripeai~lanuginosumSchltr.InFeddeRepert.Sp.Nov.Beih.4:38.1919.——Cypripediumcorr~pactumSchltr.,1.C.12:327.1922.——Cypripedi·Nnmacranthumauct.nonSw.:西藏植物志5:654.图338.1987.
亚组概述:Subsect.Macrantha(Kraenzl.)Cribb,GenusCypripedium:190.1997.---Cypripediumsect.Calceolariagroup2.MacranthaKraenzl,Orch.Gen.Sp.1:24.1901.子房具短柔毛或无毛;花1朵;花瓣一般不扭转;退化雄蕊基部通常无明显的柄。本亚组模式种:CypripediummacranthumSw.共7种,分布于中国西部至喜马拉雅地区。我国均产之。
本组概述:茎长或短,具2至多枚互生叶;叶片椭圆形、卵形至披针形,具平行脉,常被短柔毛;花单生或2—3朵排成总状花序;子房具腺毛或柔毛,较少无毛;合萼片存在;花瓣常较狭长,扭转或不扭转;退化雄蕊扁平,基部有柄或近无柄。本组模式种:CypripediumcalceolusL.共有18种,分布于北温带,以北美与东亚为最多。我国有15种。亚组1.杓兰亚组Subsect.Cypripedium子房具腺毛;花1—3朵;花瓣常扭转,较少不扭转;退化雄蕊基部通常有柄。本亚组模式种与组同。共有11个种,产北温带,主要见于东亚和北美。我国有7种。
本属概述:L,SpPl.Ed.1,2:951.1753;Pfitz.InEngler,PflanzenreidhⅣ(50)Heft12:28.1903;S.C.ChenetY.Z.XiinProc.12thWorldOrch.Conf.145.1987;Cribb,GenusCypripediuin1——301.1997.地生草本,具短或长的横走根状茎和许多较粗厚的纤维根。茎直立,长或短,成簇生长或疏离,无毛或具毛,基部常有数枚鞘。叶2至数枚,互生、近对生或对生,有时近铺地;叶片通常椭圆形至卵形,较少心形或扇形,具折扇状脉、放射状脉或3—5条主脉,有时有黑紫色斑点。花序顶生,通常具单花或少数具2—3花,极罕具5--7花;花苞片通常叶状,明显小于叶,少有非叶状或不存在;花大,通常较美丽;中萼片直立或俯倾于唇瓣之上;2枚侧萼片通常合生而成合萼片,仅先端分离,位于唇瓣下方,极罕完全离生;花瓣平展、下垂或围抱唇瓣,有时扭转;唇瓣为深囊状,球形、椭圆形或其他形状,一般有宽阔的囊口,囊口有内弯的侧裂片和前部边缘,囊内常有毛;蕊柱短,圆柱形,常下弯,具2枚侧生的能育雄蕊、1枚位于上方的退化雄蕊和1个位于下方的柱头;花药2室,具很短的花丝;花粉粉质或带粘性,但不粘合成花粉团块;退化雄蕊通常扁平,椭圆形、卵形或其他形状,有柄或无柄,极罕舌状或线形;柱头肥厚,略有不明显的3裂,表面有乳突。果实为蒴果。本属后选模式种:CypdpediumcalceolusL.全属约50种,主要产东亚、北美、欧洲等温带地区和亚热带山地,向南可达喜马拉雅地区和中美洲的危地马拉。我国有32种,广布于自东北地区至西南山地和台湾高山,绝大多数种类均可供观赏。
亚科概述:Lindl,Coil.Bot.App7.1821,“Cypripedieae”,etOrch.Scelet.7.1826;GarayinBot.Mus.Leafl.HarvardUniv.19:86.1960;DressierinSelbyana5:201.1979.地生或附生草本,极罕近亚灌木状。茎短或长,一般不分枝。叶茎生或基生,折扇状或对折。花序顶生,总状或减退为单花;子房1室或3室;花通常较大,由于子房弯曲而使唇瓣处于下方;侧萼片一般合生而成合萼片,位于唇瓣下方;唇瓣深囊状;蕊柱位于囊口,具2枚能育雄蕊、1枚退化雄蕊和1个柱头;能育雄蕊通常具短的花丝和较小的花药;花粉多少有粘性,但不形成团块;退化雄蕊位于上方,一般较大;柱头位于下方,肥厚。果实为蒴果。种子通常有翅状种皮,较少例外。本亚科模式属:CypripediumL.共含4属(近年发表的Mexipedium子房1室,似应归人Paphiopedilum),产北温带至美洲与亚洲的热带地区以及大洋洲。我国有2属。
本科概述:地生、附生或较少为腐生草本,极罕为攀援藤本;地生与腐生种类常有块茎或肥厚的根状茎,附生种类常有由茎的一部分膨大而成的肉质假鳞茎。叶基生或茎生,后者通常互生或生于假鳞茎顶端或近顶端处,扁平或有时圆柱形或两侧压扁,基部具或不具关节。花葶或花序顶生或侧生;花常排列成总状花序或圆锥花序,少有为缩短的头状花序或减退为单花,两性,通常两侧对称;花被片6,2轮;萼片离生或不同程度的合生;中央1枚花瓣的形态常有较大的特化,明显不同于2枚侧生花瓣,称唇瓣,唇瓣由于花(花梗和子房)作180°扭转或90°弯曲,常处于下方(远轴的一方);子房下位,1室,侧膜胎座,较少3室而具中轴胎座;除子房外整个雌雄蕊器官完全融合成柱状体,称蕊柱;蕊柱顶端一般具药床和1个花药,腹面有1个柱头穴,柱头与花药之间有1个舌状器官,称蕊喙(源自柱头上裂片),极罕具2—3枚花药(雄蕊)、2个隆起的柱头或不具蕊喙的;蕊柱基部有时向前下方延伸成足状,称蕊柱足,此时2枚侧萼片基部常着生于蕊柱足上,形成囊状结构,称萼囊;花粉通常粘合成团块,称花粉团,花粉团的一端常变成柄状物,称花粉团柄;花粉团柄连接于由蕊喙的一部分变成固态粘块即粘盘上,有时粘盘还有柄状附属物,称粘盘柄;花粉团、花粉团柄、粘盘柄和粘盘连接在一起,称花粉块,但有的花粉块不具花粉团柄或粘盘柄,有的不具粘盘而只有粘质团。果实通常为蒴果,较少呈荚果状,具极多种子。种子细小,无胚乳,种皮常在两端延长成翅状。科的模式属:OrchisL.全科约有700属20000种,产全球热带地区和亚热带地区,少数种类也见于温带地区。我国有171属1247种以及许多亚种、变种和变型。兰科是一个进化而复杂的科,至今对科内各类群间亲缘关系的了解仍然是十分初步的。在目前已发表的许多兰科系统中,以R.Dressier(1981,1993)系统较为全面,但该系统对鸟巢兰族(ridb.Neottieae)的处理是有欠缺的。幸而C.Seidenfaden等人在“TheOrehidsofIndochina”(OperaBot.114:5--502.1992)和“TheOrehidsofPeninsularMalaysiaandSingapore”(1992)中有较合理的处理。本书基本上采用了后者的框架,但将等级作了调整,而对亚族的组成则更多地参考Dressler系统(PhylogenyandClassifica-tionoftheOn:hidFamily,1993)。本书将兰科分为3个亚科,兰亚科之下分4个族。其中兰族(Orchideae)与万代兰族(Vandeae)是比较自然的类群,而鸟巢兰族(Neottieae)与树兰族(Epidendreae)则尚存在不少问题。在许多近代兰科系统中,上述的4个族均被作为亚科,在亚科之下又有大量的族,在族之下则是亚族。应当说,大多数亚族的建立是比较客观的。但族的组合则含有相当多的主观性,甚至是勉强的拼凑。因此,本书避繁就简,在兰亚科之下只承认4个族,族之下则包含较多的亚族。这样不仅简化了层次,而且更为客观和实用。当然,这也只是权宜之计,有些问题仍然未能解决,如鸟巢兰族与树兰族的特征交叉过多,有些亚族较为庞杂等。这里就不作详细讨论了。这里要说明的是,本书所提供的“亚科与族的检索表”和“兰亚科分亚族检索表”均系根据国产种类编写而成的,未必完全符合族与亚族的全面特征,在使用时务请加以注意。
亚科概述:Lindl,Coil.Bot.App7.1821,“Cypripedieae”,etOrch.Scelet.7.1826;GarayinBot.Mus.Leafl.HarvardUniv.19:86.1960;DressierinSelbyana5:201.1979.地生或附生草本,极罕近亚灌木状。茎短或长,一般不分枝。叶茎生或基生,折扇状或对折。花序顶生,总状或减退为单花;子房1室或3室;花通常较大,由于子房弯曲而使唇瓣处于下方;侧萼片一般合生而成合萼片,位于唇瓣下方;唇瓣深囊状;蕊柱位于囊口,具2枚能育雄蕊、1枚退化雄蕊和1个柱头;能育雄蕊通常具短的花丝和较小的花药;花粉多少有粘性,但不形成团块;退化雄蕊位于上方,一般较大;柱头位于下方,肥厚。果实为蒴果。种子通常有翅状种皮,较少例外。本亚科模式属:CypripediumL.共含4属(近年发表的Mexipedium子房1室,似应归人Paphiopedilum),产北温带至美洲与亚洲的热带地区以及大洋洲。我国有2属。
本科概述:地生、附生或较少为腐生草本,极罕为攀援藤本;地生与腐生种类常有块茎或肥厚的根状茎,附生种类常有由茎的一部分膨大而成的肉质假鳞茎。叶基生或茎生,后者通常互生或生于假鳞茎顶端或近顶端处,扁平或有时圆柱形或两侧压扁,基部具或不具关节。花葶或花序顶生或侧生;花常排列成总状花序或圆锥花序,少有为缩短的头状花序或减退为单花,两性,通常两侧对称;花被片6,2轮;萼片离生或不同程度的合生;中央1枚花瓣的形态常有较大的特化,明显不同于2枚侧生花瓣,称唇瓣,唇瓣由于花(花梗和子房)作180°扭转或90°弯曲,常处于下方(远轴的一方);子房下位,1室,侧膜胎座,较少3室而具中轴胎座;除子房外整个雌雄蕊器官完全融合成柱状体,称蕊柱;蕊柱顶端一般具药床和1个花药,腹面有1个柱头穴,柱头与花药之间有1个舌状器官,称蕊喙(源自柱头上裂片),极罕具2—3枚花药(雄蕊)、2个隆起的柱头或不具蕊喙的;蕊柱基部有时向前下方延伸成足状,称蕊柱足,此时2枚侧萼片基部常着生于蕊柱足上,形成囊状结构,称萼囊;花粉通常粘合成团块,称花粉团,花粉团的一端常变成柄状物,称花粉团柄;花粉团柄连接于由蕊喙的一部分变成固态粘块即粘盘上,有时粘盘还有柄状附属物,称粘盘柄;花粉团、花粉团柄、粘盘柄和粘盘连接在一起,称花粉块,但有的花粉块不具花粉团柄或粘盘柄,有的不具粘盘而只有粘质团。果实通常为蒴果,较少呈荚果状,具极多种子。种子细小,无胚乳,种皮常在两端延长成翅状。科的模式属:OrchisL.全科约有700属20000种,产全球热带地区和亚热带地区,少数种类也见于温带地区。我国有171属1247种以及许多亚种、变种和变型。兰科是一个进化而复杂的科,至今对科内各类群间亲缘关系的了解仍然是十分初步的。在目前已发表的许多兰科系统中,以R.Dressier(1981,1993)系统较为全面,但该系统对鸟巢兰族(ridb.Neottieae)的处理是有欠缺的。幸而C.Seidenfaden等人在“TheOrehidsofIndochina”(OperaBot.114:5--502.1992)和“TheOrehidsofPeninsularMalaysiaandSingapore”(1992)中有较合理的处理。本书基本上采用了后者的框架,但将等级作了调整,而对亚族的组成则更多地参考Dressler系统(PhylogenyandClassifica-tionoftheOn:hidFamily,1993)。本书将兰科分为3个亚科,兰亚科之下分4个族。其中兰族(Orchideae)与万代兰族(Vandeae)是比较自然的类群,而鸟巢兰族(Neottieae)与树兰族(Epidendreae)则尚存在不少问题。在许多近代兰科系统中,上述的4个族均被作为亚科,在亚科之下又有大量的族,在族之下则是亚族。应当说,大多数亚族的建立是比较客观的。但族的组合则含有相当多的主观性,甚至是勉强的拼凑。因此,本书避繁就简,在兰亚科之下只承认4个族,族之下则包含较多的亚族。这样不仅简化了层次,而且更为客观和实用。当然,这也只是权宜之计,有些问题仍然未能解决,如鸟巢兰族与树兰族的特征交叉过多,有些亚族较为庞杂等。这里就不作详细讨论了。这里要说明的是,本书所提供的“亚科与族的检索表”和“兰亚科分亚族检索表”均系根据国产种类编写而成的,未必完全符合族与亚族的全面特征,在使用时务请加以注意。 |


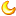
 發表於 2010-10-2 01:16 AM
|
發表於 2010-10-2 01:16 AM
| 




